
 分享
分享
1
流感嗜血杆菌(H. influenzae),氨苄西林的耐药性,可推测阿莫西林敏感情况。氨苄西林耐药主要因为产β-内酰胺酶。产β-内酰胺酶(主要是TEM-1,也可是ROB-1)的流感嗜血杆菌菌株,对氨苄西林和阿莫西林耐药,其MIC常≥128μg/ml。
2
不产β-内酰胺酶的流感嗜血杆菌,对氨苄西林耐药可由ftsI基因点突变影响青霉素结合蛋白(PBPs),β-内酰胺无法与细胞壁上作用靶点结合,PBPs亲和性降低,称为β-内酰胺酶阴性氨苄西林耐药(β-lactamase-negative and ampicillin-resistant,BLNAR),对氨基青霉素-β-内酰胺酶抑制剂类抗生素(如阿莫西林-克拉维酸、氨苄西林-舒巴坦、哌拉西林-他唑巴坦)和第一代与二代头孢类抗生素耐药。哌拉西林与哌拉西林-他唑巴坦虽然较少受到PBP介导的耐药机制的影响,但其尚无证据表明其对BLNAR的流感嗜血杆菌有临床疗效。
3
流感嗜血杆菌分离株的PBPs改变和产β-内酰胺酶的情况越来越多见。这些菌株对于阿莫西林-克拉维酸与氨苄西林-舒巴坦耐药(β-内酰胺酶阳性,对阿莫西林-克拉维酸耐药,就视作对哌拉西林-他唑巴坦、一二代头孢均耐药)。
4
目前未见产ESBL的流感嗜血杆菌,但是曾检出blaESBL基因,如同时伴有PBP3改变,将致其对三代头孢耐药。此外,在副流感嗜血杆菌中曾检出TEM ESBL变体。
流感嗜血杆菌药敏报告可参照表格:
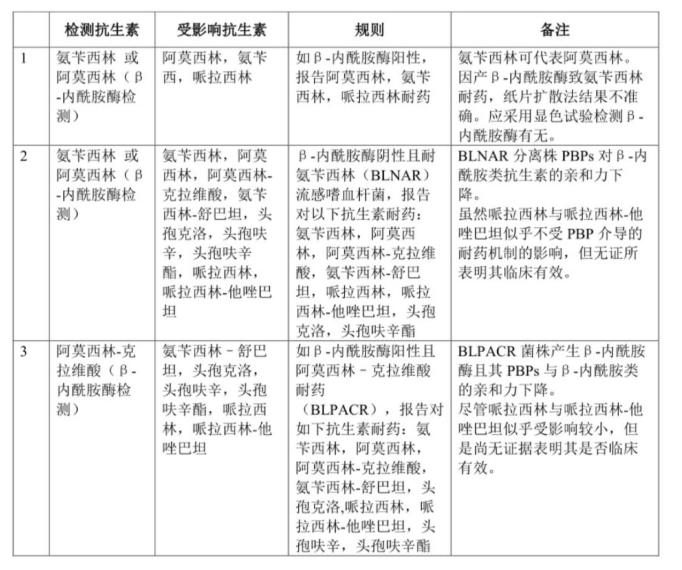
流感嗜血杆菌耐药机制与药敏结果解释
5
碳青霉烯类抗生素对于流感嗜血杆菌的不敏感情况罕见,偶有报道,多为PBP3结构改变所致。此外,也可能是抗生素进入菌内速度下降或外排机制变异所致 [1]。
6
脑脊液的分离株,应出具氨苄西林、一种三代头孢(如头孢噻肟、头孢他啶、头孢唑肟、头孢曲松)、美洛培南药敏结果 [2]。
7
药敏试验时,配制0.5号麦氏单位菌悬液相当于1-4×10^8CFU/mL。配制浓度过高会导致对某些β内酰胺类抗生素假性耐药结果,尤其是β-内酰胺酶的菌株 [2]。
8
流感嗜血杆菌对于任一三代头孢、碳青霉烯类及喹诺酮类抗生素耐药的情况,非常罕见,报告结果之前,需要复核。
参考文献
[1] Kitaoka K, Kimura K, Kitanaka H, Banno H, Jin W, Wachino JI, Arakawa Y: Carbapenem-non-Susceptible Haemophilus influenzae with Penicillin-Binding Protein 3 containing Amino Acid Insertion. Antimicrobial agents and chemotherapy 2018.
[2] CLSI.: Performance Standards for Antimicrobial Susceptibility Testing; 27nd Informational Supplement M100-A27. Wayne, PA: Clinical and Laboratory Standards Institute. 2017.
[3] Leclercq R, Canton R, Brown DF, Giske CG, Heisig P, MacGowan AP, Mouton JW, Nordmann P, Rodloff AC, Rossolini GM et al: EUCAST expert rules in antimicrobial susceptibility testing. Clinical microbiology and infection : the official publication of the European Society of Clinical Microbiology and Infectious Diseases 2013, 19(2):141-160.
作者介绍

本文转载自订阅号:京港感染论坛(ID:PIDMIC)